Below the asterisks I've copied some remarks about this disorder from my article, First Person: Neuro-Cognitive Notes on the Self in Life and in Fiction, PSYART: A Hyperlink Journal for Psychological Study of the Arts, August 21, 2000.
* * * * *
There is another aspect of the neural self, one that has to do with the continuity and coherence of the representation. We can approach this issue by considering dissociative identity disorder (DID), an extreme pathology in which the neural self is fractured. In DID, also known as multiple personality disorder, one biological individual exhibits several different identities, each having different memories and personal style. In Thigpen and Cleckley's (1957) classic study Eve had three personalities; Schreiber's (1973) Sybil had sixteen (see also Rappaport 1971, Stoller 1973). Although there has been some controversy over whether or not DID is real or simply the effect of zealous therapeutic invention and intervention, there is no doubt that at least some cases are genuine (Schachter 1996, 236-242, Spiegel 1995, 135-138).
These different identities have different personal histories. The events in one personal history typically are unknown to the other histories. Each identity will have blank periods in its history, intervals, obviously, where another identity was being enacted. And the different "persons" are often unaware of one another. Further, the different identities seem to have different personal styles, different modes of speech, of movement, of dress, and so forth. Thus both the core and autobiographical selves seem to be riven. Using the conventions we employed above, Figure 7 is a simple depiction of DID:
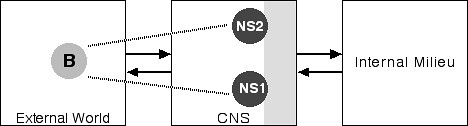
Figure 7: Dissociative Identity Disorder
Notice that we now have two neural selves, NS1 and NS2, corresponding to two different identities. Of course, these two selves exist in the same body, so we have only one corresponding body in the external world.
We do not, so far as I know, understand why or how DID happens. It is not, however, the result of the sort of gross destruction of brain tissue that underlies anosognosia. One might imagine, for example, that the different selves reside in distinctly different patches of neural tissue, a speculation that Damasio (1999b, 355) himself has suggested for the autobiographical self (though he presents no evidence). This suggestion, however, has at least one problem: How does the nervous system switch from one identity to another? There is another way of explaining DID, equally speculative and equally without specific evidence, that eliminates this particular problem.
Noting that different the identities seem to favor different moods and that "memories established in one mood state are often more readily recalled in that same mood state than in a different one," Daniel Schachter (1996, 238) suggests that "different moods and roles come to be labeled with separate names" (cf., LeDoux 1996, 211-212, McGaugh 1995, Cahill and McGaugh 1996). Different selves emerge to handle different desires and emotions. This suggests problems with brain neurochemistry; for our moods, emotions, and desires are subserved by complex chemical interactions in the nervous system (Bremmer et al. 1996, Freeman 1995, 117 ff.; Joseph 1999, Panksepp 1998, 1999). The distribution of many of these neurochemicals is regulated by brain stem nucleii, at least some of which are in the system which Damasio (1999a, 1999b) calls the proto-self – also includes cortical structures. While the activities of the core and autobiographical selves are conscious, those of the proto-self are not (though others have a different view of the structures of this proto-self, e.g. Panksepp 1998, 309 ff.). However, the structures of the proto-self help constitute the conscious states in which the core and autobiographical selves operate (cf. Hobson 1999a, 1999b).
Among these brain stem nucleii are those of the reticular formation (RF), one of the oldest structures in the brain. Classically (Moruzzi and Magoun 1949) the reticular formation has been associated with sleep and arousal. However, others have argued that the RF plays a broader role. Thus Vanderwolf and Robinson (1981) have argued that the RF exerts a general role in the control of adaptive behavior through its ability to influence the cortex. More recently, Damasio has argued that the RF and closely associated structures play a critical role in "managing body states and representing current body states. Those activities are not incidental to the brain stem's well-established activation role: "they may be the reason why such an activation role has been maintained evolutionarily and why it is primarily operated from that region" (Damasio 1999, 274).
These views are reminiscent of a very elegant model proposed by William Kilmer and Warren McCulloch, one of the "first models of decision making in neural circuitry to explicitly opt for cooperative computation, rather than executive control" (Amari and Arbib, 1977, 119). Noting that "No animal can, for instance, fight, go to sleep, run away, and make love all at once" Kilmer-McCulloch went on to list fifteen "mutually incompatible modes of vertebrate behavior," all of them as basic as those already mentioned (Kilmer, McCulloch, and Blum 1969, 279). The exact number and identity of these modes is not important. What is important is that, at all times, an animal must be in one of these modes, and only one of them. Kilmer-McCulloch hypothesized that it was the RF that determined which mode the animal was committed to at any moment (cf. Benzon and Hays 1988, pp. 296-298). The RF has extensive afferent connections from the rest of the brain and the structure of its internal connections seems well-suited to making a global evaluation of those inputs, thereby assessing the current state of the organism. The RF also has extensive efferent connections to the rest of the brain and is thus in a position to affect its state (cf. Panksepp 1999, 21 ff.; Green 1999, 43). Specifically, the RF is in a position to affect the neurochemical ambiance of cortical tissue and thus can affect just which synapses are most arousable at any given moment.
In this view, if the animal is in eating mode, the world becomes an array of objects and events that either promise something to eat, or interfere with that promise. The cortical patterns most relevant to eating are readily arousable while those less relevant are less arousable. Similarly, if the animal is in mating mode, then the world becomes a set of opportunities for sexual satisfaction or frustration. The RF commits the brain to a pattern of activation that is suitable to the mode and the animal then seeks its satisfaction. Just what it does is not directly determined by the RF; that depends on other neural centers. Figure 8 is a highly schematic representation of mode:
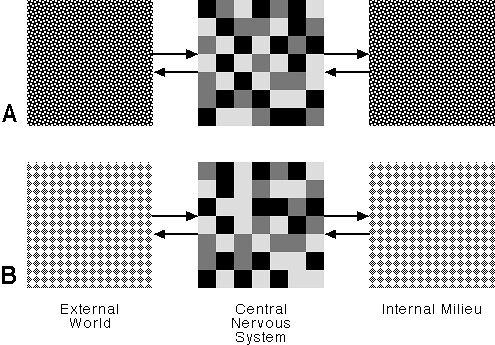
Figure 8: Behavioral Mode
Here we see two different modes, A and B. I've used some arbitrary pattern to color the internal milieu and external world for each mode, thus indicating that the animal is committed to bringing about a certain kind of consonance between its inner milieu and the external world. I've depicted the modal arousal of the CNS by varying patterns of shaded squares, where the shading corresponds to level of arousal. I've used three levels of arousal, but there could be only two, there could be seven, or level of arousal could be continuously variable. For our immediate purposes it makes no difference. What's important is that we have two distinctly different patterns of arousal corresponding to two behavioral modes.
My suggestion about DID, then, is that the mechanism that switches between one identity and another is fundamentally neurochemical. Each identity favors a particular mode, or, more likely, a set of modes. An identity becomes regnant when brain neurochemistry favors it. The perceptions and memories relevant to the modes of that identity will become easily arousable while those relevant to other modes will be all but impossible to arouse. Among individuals unaffected by DID the neurochemical milieu will bias cortical tissue toward a particular set of perceptions and memories but will not necessarily make other perceptions and memories impossible to reconstruct. In the case of DID this neurochemical process is taken to an extreme where whole ranges of perceptions and memories become absolutely unavailable depending on what neurochemicals are currently active. The state space of the brain has become fractured along neurochemical lines, breaking the self into many selves [Note 1].
Note that this explanation of DID not only tells us how the brain switches from one identity to another – RF control over cortical arousal – but also suggests that the various neural selves do not have to be in physically separate tissue. They can exist within the same volume of neural tissue for they are differentiated by chemical sensitivity, not by location.
No comments:
Post a Comment