Yesterday I introduced the concept of behavioral mode through a discussion of Shakespeare’s Sonnet 129. Now I want to present a more theoretical discussion, one which is rewritten from a paper David Hays and I published over two decades ago: Principles and Development of Natural Intelligence, Journal of Social and Biological Structures, 1988, pp. 293-322. To facilitate presentation I’ve stripped out almost all of the citations, but you can find them in the original paper.
Our object in that was to get some basic theoretical “purchase” on the brain. To that end we reviewed a wide range of observations and thinking in cognitive and neuroscience, developmental biology, and comparative psychology and arrived a five principles we thought of as governing the integration of perceptual and cognitive operations. We argued that the principles had an intrinsic ordering “such that implementation of each principle presupposes the prior implementation of its predecessor.” The modal principle is the most basic one and we regarded it as intrinsic to all vertebrate nervous systems.
Our explication began with a simple observation: the nervous system operates in two environments, an inner and an external one.

Figure 1: Action in Two Environments
It must direct the organism to act in the external environment so as to meet demands set and sensed in the inner environment; this activity is mediated by the external senses and by the skeletal muscle system. The nervous system also acts on the inner environment, sometimes directly through immediate control over respiration, heart beat, etc. and sometimes indirectly through its control of the endocrine system. Control over behavioral mode is the nervous system's basic means of coordinating activity in inner and outer environments. The brain is part of the inner environment.
The concept of modal control was originally explicated by Kilmer, McCulloch & Blum (1969) in an account of the reticular formation. They argued that animals must always be in one of several mutually exclusive modes of behavior and that the reticular formation, with its extensive afferent and efferent connections to the rest of the nervous system, is the obvious structure for implementing that commitment. The reticular formation facilitates activity in those brain regions which are most important for the current mode, while the actual behavior of the organism when it is in the mode will be regulated by other brain centers and systems.
Thus, if the animal is in eating mode, the reticular system will keep that mode active until the animal’s hunger is satisfied. But just what the animal will eat, and how it will eat it, that’s not determined by the reticular system, which ‘knows’ nothing of such things. The execution of the behavior will be handled by other, ‘higher,’ systems.
Figure 2 presents this in a highly schematic way. Here we see the organism in two situations, A and B. The external physical environment may well be the same in each case, but the organism’s priorities are different, hence a different pattern of activation in the central nervous system (center column).
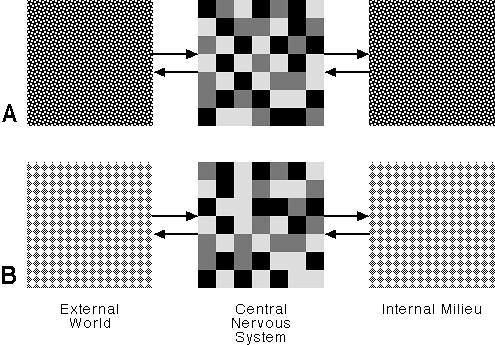
Figure 2: Two Behavioral Modes
Kilmer et al. list 15 different modes, including, for example, sleeping, eating, fighting, defecation, hunting and grooming. Whether or not McCulloch’s list is correct is beside the point; what is important is recognizing that there is some small finite list of behavioral modes. McCulloch conceived the basic reticular ‘calculation’ as being competitive. The reticular system receives inputs from all over the brain and evaluates their significance. Each mode is, in effect, attempting to ‘capture’ neural resources in service of its particular objective by interpreting its inputs as evidence for the salience of its activity, whether sleeping, eating, or grooming, etc. Only one activity can be performed at a time, hence the competitive nature of the computation.
This strategy for relating the modal principles to gross neuro-anatomy sees the brain as organized in layers about a central core. The central core, the reticular formation, is one of the oldest structures in the nervous system. It sets the mode for the next layer, and each successive layer can be seen as setting the mode for the layers peripheral to it. Modes are effected by facilitating information processing in those areas of the brain which are most relevant to the corresponding behavior and by turning control over to the appropriate neural centers. The selection of a particular behavioral goal, the formation of a plan to attain the goal and monitoring the success of that plan are all peripheral to the mode setting mechanism. Once the goal appropriate to a given mode has been attained, that mode will weigh less strongly in the reticular calculations which will then commit the brain to a different mode. The fundamental act is to commit the brain to a configuration of activity which interprets the external world as an arena for the satisfaction of particular inner needs, e.g. for nutrition, a sex partner, etc.
Biological principles are about maintaining the physical integrity of the organism, and, by implication, of the neural substrate which implements the informatic principles. Through the modal principle, the organism ultimately ensures that the schemas stored in other tissue have adaptive value. It does this through the implementation of the higher level modes of reorganization, play, imitation and speech. This account of reticular function is consistent with Vanderwolf & Robinson's (1981) review, which focuses on its role in regulating the activity of the cortex in a wide range of adaptive behaviors. The cortex implements the higher informatic functions and the reticular formation presides over their adaptive value.
The idea that differential activation of various brain areas subserves modal commitment is made particularly vivid by the various techniques which have been developed for displaying patterns of blood flow in the cortex, which show differences in patterns of neocortical activation according to such behavioral modes as voluntary movement, speaking, reading silently, reading aloud. The linguistic modes are certainly unique to man. The important point here is that they are distinct behavioral modes, requiring different configurations of brain activity.
A rather different set of findings which can fruitfully be interpreted as clues to the biochemical implementation of the modal principle is the discovery of neuropeptides whose chemical structure is identical to those of gut hormones. What is particularly interesting is that these peptides have been discovered in brain regions which are apparently unrelated to the peripheral functions mediated by the corresponding gut hormone. Thus, for example, sex hormones have been found in the sonic motor nuclei of the oyster toadfish, and in auditory and vocal nuclei of South African clawed frogs. In conjunction with evidence that neuronal local circuits are sensitive to chemical ambience, this suggests that modal control involves not only facilitating activity of various areas of the nervous system according to the demands of a given behavioral mode, but also a process of tagging or labeling stored patterns according to the modes which they serve.
The modal function, the need to co-ordinate activity in the internal and external environments in commitment to a behavioral mode, thus can provide a way of interpreting two sets of phenomena: (1) gross patterns of activation in brain regions, and (2) the prevalence of neuropeptides which are identical to gut hormones and which might thus serve to label stored patterns. These two mechanisms can be seen as the means by which biological materials implement the psychological function of mode control.
This discussion needs two qualifications. McCulloch seems to have conceived of the modal system as being flat. But how does one account for play, where a young animal ‘pretends’ to fight, or hunt, or to build a nest? These activities are themselves modal, suggesting that play is, in fact, a higher level mode. The other issue concerns such human behaviors as music-making, speech, writing, calculation, and so forth. Each has a different overall pattern of cortical activity, making them modal in the sense that Hays and I adopted from McCulloch. Does that mean that they are somehow directly regulated by the reticular system, that old old tangle of tissue at the core of the brain? That seems unlikely, especially as it would seem to imply changes in neurobiology subserving calculation and writing, for which there is no evidence. It is by no means clear to me how these modes are regulated, but will address that problem in the next post in this series.
Finally, let us recall the previous post, where the neurochemical basis of behavior was seen as causing some nasty problems. Sexual desire is one mode; consummation another; and recollection and moral evaluation still another mode. But reason does not rule that particular roost. Rather, it would seem that the reticular system exerts a strong influence on the lust cycle. Given that the reticular system is phylogenetically old and located deep in the brain’s core, that would seem to indicate that our behavior is ruled as much from the bottom as the top. That, of course, is not news to philosophers or psychoanalysts, but it may be something of a shock to all those investigators who lodge executive control in the neocortex, generally the frontal or pre-frontal lobes. Whatever that executive’s range of authority, it does not have authority over the reticular system. Rather, this executive’s capacity to command and control is itself subject to reticular activity. Which is why the executive is powerless in the face of overwhelming sexual desire or overwhelming guilt.
This suggests, in turn, that an understanding of the reticular system and of McCulloch’s concept of behavioral mode would be very useful in the neural reconstruction of psychoanalytic theory and models. Ironically enough, McCulloch himself was an intellectual foe of psychoanalysis. Is his concept of behavioral mode, then, a particularly vengeful case of the return of the repressed?
References
Kilmer, W. L., W. S. McCulloch, et al. (1969). "A Model of the Vertebrate Central Command System." International Journal of Man-Machine Studies 1: 279-309.
Vanderwolf, C. H. and T. E. Robinson (1981). "Reticulo-cortical Activity and Behavior: A Critique of the Arousal Theory and a New Synthesis." Behavioral and Brain Sciences 4: 459-576.
Addendum Oct 2014: For recent work based on McCulloch's idea, see Bernhard J. Mitterauer, Model of the Reticular Formation of the Brainstem Based on Glial–Neuronal Interactions, Cognitive Computation, June 2014. See also Michael Arbib, Modular models of brain function, in the Scholarpedia.
Addendum Oct 2014: For recent work based on McCulloch's idea, see Bernhard J. Mitterauer, Model of the Reticular Formation of the Brainstem Based on Glial–Neuronal Interactions, Cognitive Computation, June 2014. See also Michael Arbib, Modular models of brain function, in the Scholarpedia.
No comments:
Post a Comment